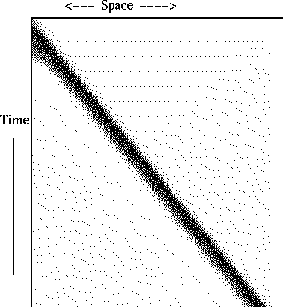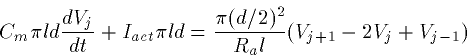
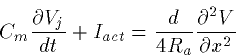
The AP provides the means of communicating between neurons by activating the axon and causing a signal to travel unattenuated down the cable. It is possible to block this by various means. For example, if the diameter of the axon increases drastically (say at a branch point) then the impuls can be blocked. Since there is no intrinsic anisotropy in the axon, it is possible to initiate an impulse any where and it will propagate outward (see Figure.) Because of the refractory period following the AP, the conducting medium behind the AP is very hyperpolarized and remains so until the sodium-potassium pump is able to rebalance the ionic concentrations. As a consequence of this, two propagating action potentials that collide, annihilate, unlike more common physical waves such as boles or light waves which pass through each other unchanged. (Note that for linear waves, this is a trivial observation, but for boles this requires some amount of mathematics.)
Repeated stimuli can have somewhat complicated effects. The
simplest is to produce a train of waves that are equally spaced dowm
the axon. If we let ![]() denote the velocity of the waves and
T their period, then there is a relationship between
denote the velocity of the waves and
T their period, then there is a relationship between
![]() and T called the dispersion relation. Any nonlinear
medium capable of producing waves has such a relationship. You have
probably observed it in water; waves with different magnitudes have
different velocities and thus tend to disperse through the medium.
Generally, the higher the temporal frequency, the smaller is the
amplitude and velocity. The fastest wave is the one with zero
frequency; the solitary pulse. The quantity,
and T called the dispersion relation. Any nonlinear
medium capable of producing waves has such a relationship. You have
probably observed it in water; waves with different magnitudes have
different velocities and thus tend to disperse through the medium.
Generally, the higher the temporal frequency, the smaller is the
amplitude and velocity. The fastest wave is the one with zero
frequency; the solitary pulse. The quantity, ![]() has
dimensions of 1/dist and is called the wavenumber or spatial
frequency of the waves. Nonmonotone dispersion relations have
profound consequences for the spacing of waves and it is possible to
get doublets and other complicated spacings of impulses. The
mathematical analysis of this type of phenomena has led to some
striking insights into propagation of the lowly action potential.
has
dimensions of 1/dist and is called the wavenumber or spatial
frequency of the waves. Nonmonotone dispersion relations have
profound consequences for the spacing of waves and it is possible to
get doublets and other complicated spacings of impulses. The
mathematical analysis of this type of phenomena has led to some
striking insights into propagation of the lowly action potential.
In spite of all this interesting behavior, when it comes time to model networks of neurons, most people ignore it and model the action potential via a conduction delay. The range of velocities are from 2-10 meters/second for a typical impulse, the faster occuring in myelinated axons. In a small piece of cortical tissue a millimeter on the side, the maximum of these delays is less than a millisecond so that for purposes of modeling we will generally neglect them. However, in parts of the olfactory cortex, there appear to be instances where conduction delays are important.