

Next: An aside on experimental
Up: Channeling with Bard
Previous: Channels - their effects
The basis for the action potential is
the voltage gated ion channel. That is the conductance of the channel
is dependent on the membrane potential at the time. This throws a
monkey wrench into the equilibrium potential since the conducatnces
are actually nonlinear function of the voltage. As in Kandel and
Schwartz, one can measure the properties of the action potential by
voltage clamp studies in order to understand the basic dynamics of
active channels. The idea is really just an application of Ohms law
and we can thus write:
| 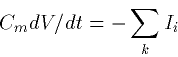 |
(2) |
where Ii are just the different currents per unit area due to the channels. Each
of these can be decomposed as:
There are at least 4 different types of channels:
- 1.
- Passive
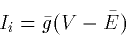
- 2.
- Persistent or noninactivating
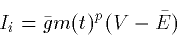
- 3.
- Transient or inactivating
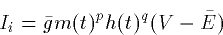
- 4.
- Anomalous or activated by hyperpolarization
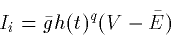
where m,h are dynamic variables that are in [0,1] and are
generally voltage dependent. m will generally increase as the
voltage increases and h decreases as the voltage increases. The
powers, p,q are meant to represent the components of a channel. For
example, the potassium channel consistes of 4 subunits and so the power
is 4.
The diagram in Figure 2 should make this
clear.
Figure 2:
The four different types of gated channels. (1) The passive
gate lets ions through a a rate proportional to the voltage drop, (2)
The persistent gate requires the ``m'' activating gate to be open, (3)
the transient gate requires both the activating gate and the
inactivating gate to be open, (4) the anomalous requires the
inactivating gate to be open. The functions 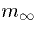 and
and 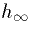 are shown in the figure
are shown in the figure
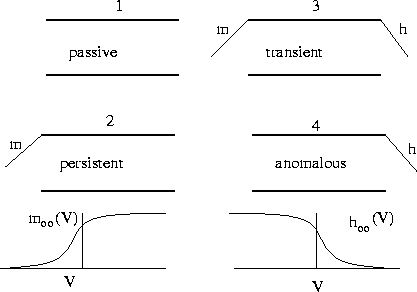 |
In the squid axon, potassium is of type 2 (persistent noninactivating)
and sodium is of type 3 (transient or inactivating.) In the thalamus,
there is a type 3 calcium current that is very important for
synchronizing spindle activity. Now lets examine the dynamics of the
channel variables, m,h. The ideas are grounded in chemical kinetics
and can be neatly summarized by the mass action model:
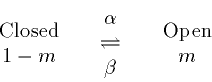
Using the law of mass action we get
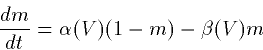
which we can rearrage to the better known form:
| 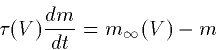 |
(3) |
where
The key points to note in general are that (i) 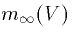 is an
increasing function of voltage for activating gates. That is,
the higher the potential, the higer the probability of the activation
gate being open. Figure 2 shows the typical activation function. (ii)
Inactivation gates decrease with increased voltage and are generally
slower than the activation gates. (iii) While the gates are generally
dependent on voltage, some potassium gates depend on the intracellular
calcium concentration or on other modulatory substances.
is an
increasing function of voltage for activating gates. That is,
the higher the potential, the higer the probability of the activation
gate being open. Figure 2 shows the typical activation function. (ii)
Inactivation gates decrease with increased voltage and are generally
slower than the activation gates. (iii) While the gates are generally
dependent on voltage, some potassium gates depend on the intracellular
calcium concentration or on other modulatory substances.
Next: An aside on experimental
Up: Channeling with Bard
Previous: Channels - their effects
G. Bard Ermentrout
1/29/1998